
Welcome everyone to my new blog. This blog is planned to be mostly about my current scientific projects and to communicate my research with other paleofolks, laypeople and colleagues alike. Maybe I will also use this platform to write about some other subjects of paleontology and related fields, but the main focus are the scientific projects I'm involved with.
In this regard, let's already start with the topic of my first article: my very first paper handled as a leading author (Abel et al. 2020). It's actually not the first paper for me to collaborate (Sachs et al. 2019a, b), but it was indeed my first paleontological project when I started it back then in 2016. The paper is partly based on my previous Bachelor's thesis, where I described a big marine crocodile from the early Upper Jurassic of Bavaria, written under the supervision of Prof. Wolfgang Kiessling at the FAU Erlangen-Nürnberg (Germany). This project was also the first time I came in touch with my now kinda regular co-authors and friends Sven Sachs (Naturkunde-Museum Bielefeld) and Dr. Mark Young (University of Edinburgh). So, I'm very happy to finally see this one out.
Marine crocodiles of Germany
In our new paper we describe the fragmentary remains of two large individuals (SMNS 80149, PSHME PH1) of marine crocodiles (Thalattosuchia) from the early
Kimmeridgian of Southern Germany. Germany is (next to e.g. the UK, France, or Argentina) one of the hotspots for fossils of these extinct crocodile relatives. Most of the well-preserved
thalattosuchian skeletons found here are from the Toarcian black shales of Holzmaden, Dotternhausen, and Mistelgau as well as the late Upper Jurassic limestones of the Altmühl valley, Wattendorf,
and Nusplingen. Additionally, there are of course also thalattosuchian remains found in Middle Jurassic, early Upper Jurassic, and Cretaceous strata. Those are, however, often only in a
fragmentary condition. Yet, even these sparse remains can tell us a lot about thalattosuchian evolution and diversity. Not to mention that they fill in an important gap between the Toarcian and
late Upper Jurassic records in Germany.

Two examples of articulated thalattosuchians from the Jurassic of Germany.
Top: two specimens of the teleosauroid "Steneosaurus" bollensis from the early Toarcian of Holzmaden, Baden-Württemberg (Paleontological collection of Tübingen).
Bottom: the metriorhynchid Cricosaurus bambergensis from the late Kimmeridgian of Wattendorf, Bavaria (Sachs et al. 2019a).
Both of our described specimens were found in thin-bedded limestones of the Subnebrodites planula Tethyan ammonite Biozone. This corresponds to the earliest Kimmeridgian (ca. 157 million years ago). For comparison, the lithographic limestones of the Altmühl valley date from the late Kimmeridgian to the early Tithonian (ca. 152 million years). So, these fossils offer a nice glimpse into the German marine reptile fauna before the famous lagoonal systems of the Altmühl valley formed.
"Drügendorf croc" and "Berreth's jaw"
Let's take a closer look at our two specimens. The one I described for my thesis (PSHME PH1) was found by hobby spelunkers in strata of the northern Bavarian Feuerstein Formation in 1999 and is now housed in the Heimatmuseum of Ebermannstadt (Bavaria). It is the more complete of the two, but "more complete" is relative of course. PSHME PH1, or how we dubbed it "Drügendorf croc", after the provenance of its fossils close to the Bavarian village of Drügendorf, consists mainly of a disarticulated torso. Most preserved bones are dorsal vertebrae and associated ribs. But also included are one of the two sacrals, an anterior caudal vertebra, the right ilium, part of the right femur as well as several conical teeth. Luckily, this is already enough for a rough classification of the animal. Especially the distinct shapes of the ilium and sacral nicely show that the "Drügendorf croc" is a member of the thalattosuchian family Metriorhynchidae.

Remains of the "Drügendorf croc" (PSHME PH1). The limestone slabs were originally aligned in one line when found in situ, the figure shows the situation as exhibited in the Heimatmuseum Ebermannstadt. Scale bar = 1 m.
Contrarily to other thalattosuchians, or basically any other crocodylomorph, metriorhynchids evolved strong adaptations to a fully-marine lifestyle. They lost their
rigid osteoderms, their limbs turned into fins, and the end of the tail evolved to form a fluke. A living metriorhynchid would not really resemble a recent crocodile in appearance, but more like
a kind of reptilian dolphin-lookalike. In this aspect, they're quite similar to the equally extinct ichthyosaurs and mosasaurs.
We can even go further in the classification of the "Drügendorf croc". The preserved teeth show on both sides sharp carinae equipped with irregular rows of
microscopic saw teeth, called denticles. Teeth with denticles typically occur in the metriorhynchid subfamily Geosaurinae. The resemblance of these teeth with saws or steak knifes is not without
reason. All known geosaurines were fairly large macropredators, adapted to feed on big prey like sharks or other marine reptiles. Conical teeth with serrated carinae are perfect for this task.
Another characteristic of the teeth in the "Drügendorf croc" are longitudinal ridges in the enamel, which are more frequent on the lingual (inner) side of the teeth and are often only restricted
to the lower half of the the tooth crowns.
This combination of tooth characters (irregular rows of microscopic denticles plus enamel ridges mostly in the lower half of the crown) is so far unknown for any metriorhynchid species, suggesting that the "Drügendorf croc" is indeed something new!
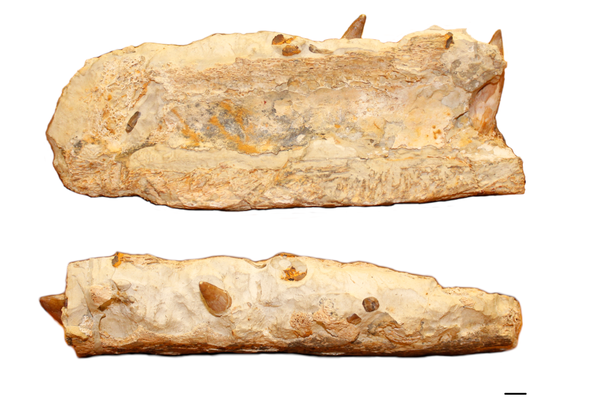
Berreth's jaw (SMNS 80149) in medial (top) and dorsal view (bottom). Scale bar = 10 mm
Our second specimen (SMNS 80149) was found by fossil collector Alfons Berreth in the 1950s or 60s in the Wohlgeschichtete Kalke Formation at Braunenberg hill near Aalen-Wasseralfingen (Baden-Württemberg), hence its nickname "Berreth's jaw". It is housed in the collection of the Museum am Löwentor Stuttgart (Baden-Württemberg). As the name implies, the fossil consists of the anterior end of a large right dentary. The dorsal surface is heavily weathered but the alveoli are still discernible with two in situ teeth preserved. While Berreth's jaw has been catalogued as a fossil of the geosaurine Dakosaurus, its teeth clearly prove otherwise.
Dakosaurus teeth are quite characteristic. They are pretty large, show no enamel ornamentation and bear relatively big regularly aligned denticles. The teeth of Berreth's jaw, however, show instead the same pattern as observed in the Drügendorf croc. Also, Berreth's jaw and Drügendorf croc derive from the earliest Kimmeridgian, contrarily to Dakosaurus, which isn't known from before the late Kimmeridgian. Together with the paleogeographic proximity and the similar paleoenvironments of the fossil-bearing strata this is a clear indication that Berreth's jaw and Drügendorf croc belonged to the same taxon, maybe even to the same species of metriorhynchids!

Comparison of different teeth of the E-clade. From left to right: Berreth's jaw, Drügendorf croc, Mr. Passmore specimen. Scale bar = 10 mm.
But where does our German taxon sit on the metriorhynchid family tree? And are there maybe other, probably unnamed metriorhynchid specimens that fit to our German fossils? As it turns out, there are.
The E-clade
Very similar teeth we also find in two other metriorhynchid fossils. These are: I) "Chouquet cf.
hastifer", an uncatalogued specimen introduced by Lepage et al. (2008) from the Kimmeridgian of France, who assign it to the enigmatic species "Metriorhynchus"
hastifer. This specimen also includes a big, nearly complete skull and several postcranial remains. And II) the so-far undescribed "Mr. Passmore specimen" (OUMNH J153).
"Berreth's jaw", "Chouquet cf. hastifer", and "Mr. Passmore specimen" further share the quite unusual shape of their alveoli, which are longitudinally distinctly shorter than in labiolingual direction. This trait again can be also observed in two other fragmentary metriorhynchid snouts from the Kimmeridgian of Switzerland and England (NMO 26589, CAMSM J18212). Lastly, in all of our specimens (apart from Drügendorf croc, where no jaws are known), are the diastemae also quite thin. The only geosaurines that show an even more extreme condition of this trait are Dakosaurus and Plesiosuchus.
Meanwhile, it could be also demonstrated that the narial openings are in our discussed specimens more retracted than in any other geosaurines (Young et al. in press). They're in this regard more similar to the highly-pelagic rhacheosaurin metriorhynchids (e.g. Cricosaurus) and other marine reptiles.
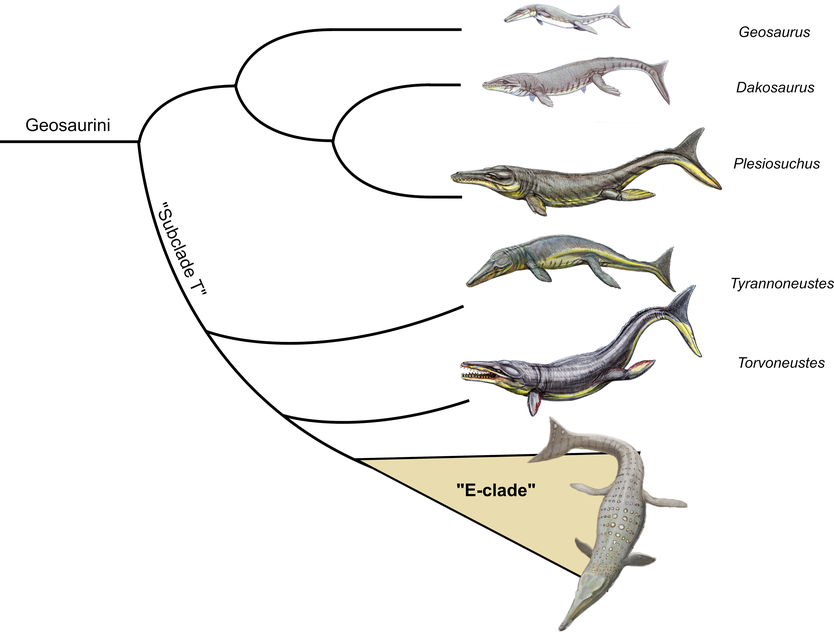
Simplified intrarelationships of geosaurin metriorhynchids, emphasizing the position of the new "E-clade". Artist of the E-clade taxon: Joschua Knüppe. All other taxa by Dmitry Bogdanov under Creative Commons licenses 2.5 and 4.0. Animals roughly to scale.
In summary, Drügendorf croc and Berreth's jaw belong to an assemblage of geosaurine metriorhynchids from the Late Jurassic of Europe that share several common traits not found in any other geosaurine. These include several characteristics of their dentition as well as the slight retraction of the narial openings. Our conducted phylogenetic analyses also support a close relationship between these specimens within one single clade, which we preliminarily call "E-clade". The E-clade itself was further found to lie within another not officially named geosaurine lineage dubbed "Subclade T", which also includes the genera Tyrannoneustes, Purranisaurus, and Torvoneustes, The specimens of the E-clade seem to be closer related to Purranisaurus and Torvoneustes than to Tyrannoneustes, but due to the very fragmentary nature of their fossils this should been taken with a grain of salt.
So, how many species are there currently in our E-clade? It is really hard to tell. I have the suspicion that both German specimens and two of the others could belong all to the same species, whereas the remaining two show slight differences to the rest. As often, more and better preserved fossils are needed to solve this.
Paleoecology and taphonomy
A last section aside to all the anatomical terminology in the previous parts. Let's talk about the actual animals behind the E-clade and also some interesting
phenomenon regarding the preservation of the Drügendorf croc. I stated already that the members of the E-clade were comparatively large metriorhynchids. But how large?
The size of the skull and the femurs proved to be good proxies to estimate the size even in incompletely preserved metriorhynchids (Young et al. 2011). Unfortunately, Drügendorf croc and Berreth's jaw are not suited for this. Nevertheless, we have the nearly complete skull of "Chouquet cf. hastifer". Measuring roughly one meter the skull is already pretty big. Our co-author Mark and his colleagues estimated the whole animal in their 2011 study to be roughly 5.4 meters long. By that it is in the same size range as other big geosaurines like Tyrannoneustes or Dakosaurus. For comparison, the currently largest known metriorhynchid is Plesiosuchus manselli with an estimated length of 6.8 meters.
And even if we don't have the necessary bones (like it's the case in the Drügendorf croc), a simple comparison of its remains with other more complete
metriorhynchids demonstrates that it was also quite an impressive individual. A length of at least 5 meters appears reasonable to me. So, the species of the new E-clade were in the upper size
range of geosaurines and comparable in size to the largest extant crocodiles.

Life restoration of the "Drügendorf croc" in the early Kimmeridigian of what is now northern Bavaria. The paleoenvironment was a life-friendly offshore region with abundant sponge-microbial reefs. The skull is based on Torvoneustes and "Chouquet cf. hastifer". Artist: Joschua Knüppe.
What did they eat? There is no hard data at the moment, but based on their skull and dental morphology, we can assume that all known
geosaurines were macropredators, however their actual diet and feeding strategy likely differed as evident by the morphological differences in their jaw apparatus. These differences also enabled
related geosaurine species and other thalattosuchians to coexist in the same environment like we can observe it for the Late Jurassic lagoons of Southern Germany (De Andrade & Young 2008). In
case of the E-clade crocs, it is for example conspicuous that both the tooth crowns and the denticles are distinctly smaller than in similar sized individuals of Dakosaurus and
Plesiosuchus. Maybe they fed on smaller prey or occasionally skimmed the richly inhabited seafloor like in the reconstruction above?
Speaking of the seafloor, the Drügendorf croc finally also offers another evidence for an interesting geological phenomenon. Carbonate sediments like the ones from the Upper Jurassic of Southern Germany often show a distinct alternation of limestone and marl layers. An intuitive interpretation of these alternations are cyclical changes in sediment input, for example due to climate changes. However, there is now more and more evidence that a number of such limestone-marl-alternations actually formed diagenetically from a mostly homogeneous carbonate sediment (e.g. Westphal et al. 2008; Nohl & Munnecke 2019).
The preservation of the Drügendorf croc is interesting in this regard, because two of its vertebrae were reportedly found in the overlying marl layer in contrast to the rest of the fossil in the limestone bed beneath. As there are no sedimentological indications for a postdepostiontal reworking of the bones or signs for a longer exposure of remains above the seafloor, a more likely explanation could be a deposition within a homogeneous carbonate sediment, with a separation of the fossil into a limestone and marl facies being a purely diagenetic product.
References:
- Abel, P., Sachs, S., Young, M.T. (2020). Metriorhynchid crocodylomorphs from the lower Kimmeridgian of Southern Germany: evidence for a new large-bodied geosaurin lineage in Europe. Alcheringa: An Australasian Journal of Palaeontology. doi: 10.1080/03115518.2019.1701079
- De Andrade, M. B., & Young, M. T. (2008). High diversity of thalattosuchian crocodylians and the niche partition in Solnhofen Sea. In: The 56th Symposium of Vertebrate Palaeontology and Comparative Anatomy, 3rd-5th September, Dublin, Dyke, G., Naish, D. and M. Parkes (Eds): 14-15.
- Lepage, Y., Buffetaut, E., Hua, S., Martin, J. E. & Tabouelle, J. (2008). Catalogue descriptif, Anatomique, Geologique et Historique des Fossiles presentes a l’Exposition “les Crocodiliens Fossiles de Normandie. Bulletin de la Societe Geologique de Normandie et des Amis du Museum du Havre 95, 5–29.
-
Nohl, T., & Munnecke, A. (2019). Reconstructing time and diagenesis of limestone-marl alternations from the selective compaction of colonies of the tabulate
coral Halysites. Bulletin of Geosciences, 94(3), 279-298. doi:
10.3140/bull.geosci.1752 - Sachs, S., Johnson, M.M., Young, M.T., and Abel, P. (2019b). The mystery of Mystriosaurus: Redescribing the poorly known Early Jurassic teleosauroid thalattosuchians Mystriosaurus laurillardi and Steneosaurus brevior. Acta Palaeontologica Polonica 64 (3): 565–579. doi: 10.4202/app.00557.2018
- Sachs, S., Young, M.T., Abel, P., and Mallison, H. (2019a). A new species of the metriorhynchid crocodylomorph Cricosaurus from the Upper Jurassic of southern Germany. Acta Palaeontologica Polonica 64 (2): 343–356. doi: 10.4202/app.00541.2018
- Westphal, H., Munnecke, A., Böhm, F., & Bornholdt, S. (2008). Limestone–marl alternations in epeiric sea settings–witnesses of environmental changes or diagnesis?. Special Paper–Geological Association of Canada, 389, 406.
- Young, M. T., Bell, M. A., De Andrade, M. B., & Brusatte, S. L. (2011). Body size estimation and evolution in metriorhynchid crocodylomorphs: implications for species diversification and niche partitioning. Zoological Journal of the Linnean Society, 163(4), 1199-1216.
- Young, M.T., Sachs, S., Abel, P., Foffa, D., Herrera, Y., Kitson, J.J.N. (in press). Convergent evolution and possible constraint in the posterodorsal retractions of the external nares in marine crocodylomorphs. Zoological Journal of the Linnean Society.
Write a comment